
Проект Академии Технологий НПО АТ
Россия — в перспективе научных идей следующего десятилетия. Что станет обязательным и актуальным? Полет человека в дальний космос и его освоение и воскрешение человека, - многовековые мечты человечества, которые станут целью стратегически крупных направлений научно-практических разработок так же официально, как сейчас поиск внеземного разума для оборонно-космической отрасли.
НПО АТ - научный проект. Мы занимаемся поиском путей воскрешения человека и открытия полета в дальний космос. Поиском и сбором данных, которые помогут человечеству идти вперед в этих направлениях. Поиском совершенно неисследованных, но перспективных для ученых мест и областей науки, которые могут дать перспективные открытия в этих направлениях. Созданием архивов данных для тех, кто хотела бы делать технические и технологические открытия и изобретения. Экономикой и организацией сложных технических лабораторий, которые делают уникальные технологические разработки в области работы в области поиска пути воскрешения человека и открытия перелета в дальний космос.
Уважаемые господа, сайт находится в разработке, приносим свои извинения за технические неполадки.
Основа приборостроения - измерение и работа биологических строительных блоков |
Биологические строительные наноблоки, их составляющие (аминокислоты) и наноструктуры
Биологические строительные наноблоки, нуклеотиды, - часть, из которых строится ДНК. Белки́ (протеи́ны, полипепти́ды) — высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной связью . В природе существует примерно 1010-1012 различных белков, обеспечивающих жизнедеятельность организмов всех степеней сложности от вирусов до человека. Белками являются ферменты, антитела, многие гормоны и другие биологические активные вещества. В живых организмах аминокислотный состав белков определяется генетическим кодом, при синтезе в большинстве случаев используется 20 стандартных аминокислот. Кроме того, аминокислотные остатки в составе белка часто подвергаются посттрансляционным модификациям, которые могут возникать и до того, как белок начинает выполнять свою функцию, и во время его «работы» в клетке. Часто в живых организмах несколько молекул разных белков образуют сложные комплексы, например, фотосинтетический комплекс.
|
Аминокислота |
Латинское название |
Химическая и эмпирическая формула |
Год открытия |
Mw, г/моль |
Размер, d ,нм |
Плотность, г/см³ |
Температура плавления, °C |
Химические свойства |
Кодируется генами (Кодо́н (кодирующий тринуклеотид) ) |
Источник |
Кто впервые выделил[1] |
|
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
аминоуксусная кислота, от др.-греч. γλυκύς, glycys — сладкий |
Glycine |
Gly, G |
NH2 – CH2 – COOH C2H5NO2 |
1820 |
75,07 |
0,42 |
1,61 |
290 Удельная теплота испарения −528,6 Дж/кг Удельная теплота плавления −981,1 Дж/кг |
pKa
2,34 |
GLRA1, GLRA2, GLRA3 и GLRB |
Желатин |
|
|
(сокр. Leu или L; 2-амино-4-метилпентановая кислота) (от «leukos» (греч.) — «белый») |
Leucine |
Leu, L |
HO2CCH(NH2)CH2CH(CH3)2 C6H13N1O2 |
1820 |
131,18 |
|
|
|
|
UUA, UUG, CUU, CUC, CUA, и CUG |
Мышечные волокна |
|
|
(α-амино-β-(п-гидроксифенил)пропионовая кислота, сокр.: Тир, Tyr, Y) — ароматическая альфа-аминокислота |
tYrosine |
Tyr, Y |
C9H11NO3 |
1848 |
181,19 а. е. м. |
|
1,46 |
343 °C |
pKa
2,24 |
Казеин |
Ф. Бопп |
|
|
Серин (α-амино-β-оксипропионовая кислота; 2-амино-3-гидроксипропановая кислота) — гидроксиаминокислота, существует в виде двух оптических изомеров — L и D |
Serine |
Ser, S |
HO2C-CH(NH2)CH2OH |
1865 |
105,09 г/моль |
|
1,537 г/см³ |
228 °C |
pKa
2,13 |
Шёлк |
Э. Крамер |
|
|
Глутаминовая кислота,(2-аминопентандиовая кислота) — алифатическая аминокислота |
gluEtamic acid |
Glu, E |
C5H9N1O4 C 40,82 %, H 6,17 %, N 9,52 %, O 43,5 % |
1866 |
147,1293 ± 0,006 |
|
1,4601 |
160 °C, Температура кипения 205 °C Температура разложения свыше 205 °C |
pKa 2,16, 4,15, 9,58 Растворимость в воде 8,64 г/100 мл Изоэлектрическая точка 3,22 |
Растительные белки |
Г. Риттхаузен |
|
|
Глутамин, 2-аминопентанамид-5-овая кислота |
Q-tamine |
Глн, Gln, Q |
C5H10N2O3 |
|
146,14 |
|
|
|
|
|
|
|
|
asparDic acid |
Асп, Asp, D |
HO2CCH(NH2)CH2CO2H C4H7NO4 |
1868 |
133,1 |
|
|
|
|
Конглутин, легумин (ростки спаржи) |
Г. Риттхаузен |
||
|
Аспарагин, (принятые сокращения: Асн, Asn, N) — амид аспарагиновой кислоты (2-амино-бутанамид-4-овая кислота, Asx или B) |
asparagiNe |
Асн, Asn, N |
HOOC-CH(NH2)-CH2-CONH2 C4H8N2O3 |
1806 |
132,12 а. е. м. |
|
|
220 моногидрат, 233 L, 182 LD °C |
pKa 2,02 и 8,84 Растворимость в воде растворим в воде Вращение [α]D + 21° Изоэлектрическая точка 5,41 |
Сок спаржи |
Л.-Н. Воклен и П. Ж. Робике |
|
|
Фенилаланин,
Анилин, Фениламин |
Fenylalanine |
Phe, F |
1881 |
93,13 г/моль |
|
1,0217 г/см³ |
Температура плавления −6,3 °C Температура кипения 184,13 °C |
Растворимость в воде 3,6 г/100 мл Динамическая
вязкость (ст. усл.)
3,71 Па·с |
|
Ростки люпина |
Э. Шульце, Й. Барбьери |
|
|
Аланин, (2-аминопропановая кислота) — алифатическая аминокислота, (S)-2-аминопропановая (α-аминопропионовая) кислота |
Alanine |
Ала, Ala, A |
CН3-СН(NH2)-СООН |
1888,19 |
Отн. молек. масса 89,09 а. е. м. Молярная масса 89,09 г/моль |
|
|
295-316 °C |
|
Фиброин шелка |
Т. Вейль, |
|
|
Лизин, (2,6-диаминогексановая кислота) — алифатическая аминокислота , 2,6-диаминогексановая кислота |
before L |
Лиз, Lys, K |
HO2CCH(NH2)(CH2)4NH2 C6H14N2O2 |
1889 |
146,19 г/моль |
|
|
|
|
Казеин |
Э. Дрексель |
|
|
aRginine |
Arg, R |
(NH-C(NH2)NH(CH2)3CH(NH2)-COOH) C6H14N4O2 |
1895 |
174,2 г/моль |
|
|
|
pKa 12,48 |
Вещество рога |
С. Гедин |
||
|
Гистидин,
(L-α-амино-β-имидазолилпропионовая кислота) —
гетероциклическая альфа-аминокислота,
L-2-амино-3-(1H-имидазол- |
Histidine |
Гис, His, H |
C6H9N3O2 |
1896 |
155,16 г/моль |
|
|
287 °C |
pKa
1,70 |
Стурин, гистоны |
А. Кессель, С. Гедин |
|
|
Цистеин, (α-амино-β-тиопропионовая кислота; 2-амино-3-меркаптопропановая кислота) — алифатическая серосодержащая аминокислота , α-амино-β-тиопропионовая кислота; 2-амино-3-меркаптопропановая кислота |
Cysteine |
Цис, Cys, C |
HO2CCH(NH2)CH2SH C3H7NO2S |
1899 |
121,16 г/моль |
|
|
|
|
Вещество рога |
К. Мёрнер |
|
|
Валин, (2-амино-3-метилбутановая кислота) — алифатическая α-аминокислота, (S)-2-амино-3-метилбутановая кислота, названи в честь валерианы |
Valine |
Вал, Val, V |
HO2CCH(NH2)CH(CH3)2 C5H11NO2 |
1901 |
117,15 г/моль |
|
1,230 г/см³ |
315 °C |
pKa
2,27 |
Казеин |
Э. Фишер, валин защищает миелиновую оболочку, окружающую нервные волокна в головном и спинном мозге. |
|
|
Пролин,
(пирролидин-α-карбоновая кислота) —
гетероциклическая аминокислота (точнее, иминокислота),
L-пирролидин-2- |
Proline |
Про, Pro, P |
C5H9NO2 |
1901 |
115,13 г/моль |
|
1,35—1,38 г/см³ |
221 °C |
pKa
1,99 |
Казеин |
||
|
Гидроксипролин,
(оксипролин, 4-гидроксипирролидин-α-карбоновая
кислота) нестандартная аминокислота,
отличающаяся от пролина
наличием гидроксильной
группы у одного из атомов углерода,
L-4-гидроксипирролидин-2- |
|
Hyp, 4-Hyp, HyPro, hP |
C5H9NO3 |
1902 |
131,13 г/моль |
|
|
|
|
|
Желатин |
|
|
Триптофан, (β-индолиламинопропионовая, или α-2-амино-3-(1H-индол-3-ил)пропионовая кислота, сокр.: Три, Трп, Trp, W) — ароматическая альфа-аминокислота. Существует в двух оптически изомерных формах — L и D и в виде рацемата (DL), 2-амино-3-(1H-индол-3-ил)пропионовая кислота |
tWo rings |
Три, Trp, W |
C11H12N2O2 |
1902 |
0,67 |
|
|
|
Казеин |
Ф. Гопкинс, Д. Кол |
||
|
Изолейцин, (сокращенно Ile или I; 2-амино-3-метилпентановая кислота)[1] — это алифатическая α-аминокислота |
Isoleucine |
Иле, Ile, I |
HO2CCH(NH2)CH(CH3)CH2CH3 C6H13O2N |
1904 |
131,17 г/моль |
|
|
|
|
Фибрин |
Ф. Эрлих |
|
|
Метионин, алифатическая серосодержащая α-аминокислота, 2-амино-4-(метилтио)бутановая кислота |
Methionine |
Met, M |
HO2CCH(NH2)CH2CH2SCH C5H11NO2S |
1922 |
149.21 г/моль |
|
1.340 г/см³ |
L- или D- 281 (разл.) °C |
pKa 2.28 (карбоксил) 9.21 (аминогруппа)[1]
|
|
Казеин |
|
|
Треонин,
(α-амино-β-гидроксимасляная кислота;
2-амино-3-гидроксибутановая кислота) —
гидроксиаминокислота; молекула содержит два хиральных
центра, что обусловливает существование четырёх оптических
изомеров: L- и D-треонина (3D), а также L- и D-аллотреонина
(3L), 2-амино-3- |
Threonine |
Тре, Thr, T |
HO2CCH(NH2)CH(OH)CH3 C4H9NO3 |
1925 |
119,12 г/моль |
|
|
256 °C |
pKa
2,20 |
Белки овса |
С. Шрайвер и др. |
|
|
Гидроксилизин, (5-гидрокси-L-лизин) — нестандартная аминокислота, (2S,5R)-2,6-диамино-5-гидроксигексановая кислота |
|
Hyl, hK, 5-Hyl |
C6H14N2O3 |
1925 |
162,187 г/моль |
|
|
|
|
|
Белки рыб |
С. Шрайвер и др. |
Данные о том, какими генами кодируются аминокислоты, молекулярным массам, плотности из Википедии 2013.
Секторный вариант записи кодона
Обратная таблица (указаны кодоны для каждой аминокислоты, а также старт — и стоп-кодоны)
|
Ala/A |
GCU, GCC, GCA, GCG |
Leu/L |
UUA, UUG, CUU, CUC, CUA, CUG |
|---|---|---|---|
|
Arg/R |
CGU, CGC, CGA, CGG, AGA, AGG |
Lys/K |
AAA, AAG |
|
Asn/N |
AAU, AAC |
Met/M |
AUG |
|
Asp/D |
GAU, GAC |
Phe/F |
UUU, UUC |
|
Cys/C |
UGU, UGC |
Pro/P |
CCU, CCC, CCA, CCG |
|
Gln/Q |
CAA, CAG |
Ser/S |
UCU, UCC, UCA, UCG, AGU, AGC |
|
Glu/E |
GAA, GAG |
Thr/T |
ACU, ACC, ACA, ACG |
|
Gly/G |
GGU, GGC, GGA, GGG |
Trp/W |
UGG |
|
His/H |
CAU, CAC |
Tyr/Y |
UAU, UAC |
|
Ile/I |
AUU, AUC, AUA |
Val/V |
GUU, GUC, GUA, GUG |
|
START |
AUG |
STOP |
UAG, UGA, UAA |
В процессе биосинтеза белка в полипептидную цепь включаются 20 α-аминокислот, кодируемых генетическим кодом. Помимо этих аминокислот, называемых протеиногенными, или стандартными, в некоторых белках присутствуют специфические нестандартные аминокислоты, возникающие из стандартных в процессе посттрансляционных модификаций. В последнее время к протеиногенным аминокислотам иногда причисляют трансляционно включаемые селеноцистеин (Sec, U) и пирролизин (Pyl, O). Это так называемые 21-я и 22-я аминокислоты.
Вопрос, почему именно эти 20 аминокислот стали «избранными», остаётся не решённым. Не совсем ясно, чем эти аминокислоты оказались предпочтительнее других похожих. Например, ключевым промежуточным метаболитом пути биосинтеза треонина, изолейцина и метионина является α-аминокислота гомосерин. Очевидно, что гомосерин — очень древний метаболит, но для треонина, изолейцина и метионина существуют аминоацил-тРНК-синтетазы, тРНК, а для гомосерина — нет.
Структурные формулы 20-ти протеиногенных аминокислот обычно приводят в виде так называемой таблицы протеиногенных аминокислот:

Большинство аминокислот, участвующих в обмене веществ входящих в состав белков, могут поступать с пищей или синтезироваться в организме в процессе обмена (из других аминокислот, поступающих в избытке). Они получили название заменимых аминокислот. Некоторые аминокислоты не могут синтезироваться в организме и должны поступать с пищей. Они получили название незаменимых.
Биологически важные реакции аминокислот:
Дезаминирование неокислительное — удаление NH2 группы без участия кислорода.
Дезаминирование окислительное — удаление NH2 группы с участием с участием ферментов дегидрогеназ и кофермента НАД+ или НАДФ+ . Большинство аминокислот подвергается непрямому Д.: после переаминирования с а-кетоглутаровой к-той образуется глутаминовая к-та, к-рая дезаминируется при участии глутаматдегидрогеназы. Др. типы Д., широко представленные у микроорганизмов: восстановительное, гидролитическое (Д. аминопроизводных пуринов, пиримидинов и сахаров) и внутримолекулярное (Д. Гистидина). (Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986.)
По механизму реакции различают окислительное (образуются ###-кетокислоты и аммиак), восстановительное, гидролитическое, внутримолекулярное Д. и др. (Источник: «Англо-русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд-во ВНИРО, 1995 г.)
Сопровождается замещением аминогруппы на др. группу или образованием кратной связи. Осуществляется под действием HNO2, нитрозилсерной к-ты, N2O3, N2O4, H2O, H2, а также ферментов. Наиб. универсальный метод Д. аминов - взаимод. их с HNO2. Сущность метода состоит в обработке кислых водных р-ров аминов нитритом К или Na, в результате чего образуются соли диазония (см. Диазотирование, Диазосоединения).В случае алифатич. аминов эти соли неустойчивы и разлагаются с выделением N2 и образованием карбкатиона, к-рый взаимод. с нуклеоф. агентами среды (вода, анионы и др.) или стабилизируется с образованием олефина, причем р-ция может сопровождаться изомеризацией алкильного остатка.
По такому же механизму осуществляется Д. алициклич. и гетероциклич. аминов, к-рая сопровождается изменением числа звеньев в цикле (см. Демьянова перегруппировка). Соли диазония ароматич. ряда значительно более устойчивы и разлагаются по гомолитич. и гетеролитич. механизму с заменой диазогруппы на др. функц. группу или атом галогена.
По гомолитич. механизму осуществляется, напр., разложение солей арилдиазония с образованием бифенила (см. Гомберга-Бахмана-Хея реакция), арилгалогенидов и азобензолов (см. Зандмейера реакция), меркурированных арилпроизводных (см. Несмеянова реакция), жирно-ароматич. соед. (см. Меервейна реакция) и др.
По гетеролитич. механизму из солей арилдиазония получают фенолы (при нагр. с Н 2 О), ариловые эфиры (в присут. NaNO3), арилфториды (см. Шимана реакция), ариларсоновые к-ты (см. Барта реакция). Под действием HNO2 протекает также Д. орг. производных гидразина с образованием аминов (см. Гидразина замещенные органические). Д. алифатич. и циклич. аминов при высокой т-ре под действием Н 2 сопровождается образованием предельных углеводородов и NH3 (см. Гидрогенолиз), Д. аминов нафталинового ряда под действием водных р-ров NaHSO3 происходит с образованием нафтолов. Под действием N2O4 амины превращаются в алкиловые эфиры азотной к-ты; амиды к-т под действием N2O3 превращаются в карбоновые к-ты с выделением NH3. Иногда Д. наз. также расщепление четвертичных аммониевых оснований (см. Гофмана реакции).
Д. аминокислот и нек-рых др. азотсодержащих соед. играет важную роль в жизнедеятельности животных, растений и микроорганизмов. Так, для a-аминокислот характерно ферментативное окислит. Д. с образованием NH3 и a-кето-кислот. Лит.: Органические реакции, пер. с англ., сб. 11, М., 1965, с. 167-98; Вейганд К., Хильгетаг Г., Методы эксперимента в органической химии, пер. с нем., М., 1968, с. 319-28. Г. В. Гришина. Источник: Химическая энциклопедия. — М.: Советская энциклопедия. Под ред. И. Л. Кнунянца. 1988.
Трансаминирование (переаминирование). Это основной путь биосинтеза a-аминокислот из a-оксокислот. Донором аминогруппы служит a-аминокислота, имеющаяся в клетках в достаточном количестве или избытке, а ее акцептором – a-оксокислота. Реакция протекает в присутствии фермента трансаминазы и кофермента пиридоксальфосфата. Это обратимый перенос аминогруппы из молекулы одного орг. соед. в молекулу другого. Наиб. роль П. играет в биохимии в процессах метаболизма азотистых оснований в тканях животных и растений. На р-циях такого типа основываются синтез и диссимиляция аминокислот.
Аминотрансферазы содержат в качестве кофсрмента производные витамина В 6 -пири-доксаль-5'-фосфат (ф-ла I) и пиридоксамин-5'-фосфат (II). В основе каталитич. активности пиридоксаль-5'-фосфата лежит способность его формильной группы образовывать с аминокислотами шиффовы основания, легко гидролизующиеся до пиридоксамин-5'-фосфата и a-кетокислоты. A. E. Браунштейн и M. M. Шемякин в 1953 сформулировали общий механизм П. в биол. системах, состоящий из ряда промежут. стадий. Подробно механизм П. под действием аминотрансфераз рассмотрен в ст. Аспартатаминотранс — Источник: Химическая энциклопедия. — М.: Советская энциклопедия. Под ред. И. Л. Кнунянца. 1988. Большинство природных аминокислот синтезируется в тканях путём переноса NH2-группы от глутаминовой кислоты (См. Глутаминовая кислота) — начального продукта усвоения азота — на различные оксокислоты. Обеспечивая быстрое взаимопревращение различных амино- и оксокислот, реакции П. играют важную роль в регуляции и сопряжении обмена аминокислот и углеводов, ферменты П.— Аминотрансферазы имеются во всех живых клетках. Описано свыше 55 различных аминотрансфераз, катализирующих П. всех известных природных аминокислот и ряда биогенных аминов. Коферментом аминотрансфераз является производное витамина B6 — пиридоксальфосфат, играющий роль переносчика NH2-группы (о механизме П. см. Пиридоксалевые ферменты). Резкое повышение содержания некоторых аминотрансфераз в плазме крови больных служит диагностическим признаком при поражениях печени (гепатиты), сердца (инфаркт миокарда), мышц (травмы, миодистрофические заболевания). Большая советская энциклопедия. — М.: Советская энциклопедия. 1969—1978.
Декарбоксилирование - a-аминокислоты ведет к образуют биогенные амины. Например , g-аминомасляная кислота принимает участие в обменных процессах, происходящих в головном мозге, является нейромедиатором. В медицинской практике под названием гаммалон, или аминалон, применяется при лечении нервно-психических заболеваний. Декарбоксилирование - отщепление СО2 от карбоксильной группы карбоновых кислот обычно при участии ферментов декарбоксилаз. Ферментативное Д. может быть обратимым (Д. оксалоацетата до пирувата) и необратимым (окислительное Д. аминокислот). Особое значение в клетке имеют реакции окислительного Д. пирувата (с образованием ацетил–КоА) и α–кетоглутарата (с образованием сукцинилкоэнзима А), что определяет функционирование цикла Кребса.
(Источник: «Микробиология: словарь терминов», Фирсов Н.Н., М: Дрофа, 2006 г.)
Гидроксилирование — например, ферментивное гидроксилирование фениламина, когда к кольцу присоединяется группа -ОН. Оно интересно влиянием на процессы мозга. Вагнера реакция - окисление олефинов перманганатом калия в слабощелочной среде, приводящее к образованию гликолей. В. р. используют для исследования строения непредельных органических соединений, содержащих этиленовые (несопряжённые) связи. С помощью В. р. было установлено строение некоторых терпенов (См. Терпены) (пинена, лимонена, терпинеола и терпина). Реакция открыта Е. Е. Вагнером в 1888.(Большая советская энциклопедия. — М.: Советская энциклопедия. 1969—1978.)
Ароматические: фенилаланин, тирозин, триптофан, (гистидин)
Иминокислоты: пролин
Класс I: валин, изолейцин, лейцин, цистеин, метионин, глутамат, глутамин, аргинин, тирозин, триптофан
Класс II: глицин, аланин, пролин, серин, треонин, аспартат, аспарагин, гистидин, фенилаланин
Для аминокислоты лизин существуют аминоацил-тРНК-синтетазы обоих классов.
Пути биосинтеза протеиногенных аминокислот разноплановы. Одна и та же аминокислота может образовываться разными путями. К тому же совершенно различные пути могут иметь очень похожие этапы. Тем не менее, имеют место и оправданы попытки классифицировать аминокислоты по путям их биосинтеза. Существует представление о следующих биосинтетических семействах аминокислот: аспартата, глутамата, серина, пирувата и пентоз. Не всегда конкретную аминокислоту можно однозначно отнести к определённому семейству; делаются поправки для конкретных организмов и учитывая преобладающий путь. По семействам аминокислоты обычно распределяют следующим образом:
Семейство аспартата: аспартат, аспарагин, треонин, изолейцин, метионин, лизин.
Семейство пентоз: гистидин, фенилаланин, тирозин, триптофан.
Фенилаланин, тирозин, триптофан иногда выделяют в семейство шикимата.
Заменимые
Классификация аминокислот на заменимые и незаменимые не лишена недостатков. К примеру, тирозин является заменимой аминокислотой только при условии достаточного поступления фенилаланина. Для больных фенилкетонурией тирозин становится незаменимой аминокислотой. Аргинин синтезируется в организме человека и считается заменимой аминокислотой, но в связи с некоторыми особенностями его метаболизма при определённых физиологических состояниях организма может быть приравнен к незаменимым. Гистидин также синтезируется в организме человека, но не всегда в достаточных количествах, потому должен поступать с пищей.
Биодеградация аминокислот может идти разными путями. По характеру продуктов катаболизма у животных протеиногенные аминокислоты делят на три группы: глюкогенные (при распаде дают метаболиты, не повышающие уровень кетоновых тел, способные относительно легко становиться субстратом для глюконеогенеза: пируват, α-кетоглутарат, сукцинил-KoA, фумарат, оксалоацетат), кетогенные (распадаются до ацетил-KoA и ацетоацетил-KoA, повышающие уровень кетоновых тел в крови животных и человека и преобразующиеся в первую очередь в липиды), глюко-кетогенные (при распаде образуются метаболиты обоих типов).
Глюкогенные: глицин, аланин, валин, пролин, серин, треонин, цистеин, метионин, аспартат, аспарагин, глутамат, глутамин, аргинин, гистидин.
Глюко-кетогенные (смешанные): изолейцин, фенилаланин, тирозин, триптофан.
Основная статья: Эксперимент Миллера — Юри
«Миллеровские» аминокислоты — обобщенное название аминокислот, получающихся в условиях, близких к эксперименту Стенли Л. Миллера 1953 года. Установлено образование в виде рацемата множества различных аминокислот, в том числе: глицин, аланин, валин, изолейцин, лейцин, пролин, серин, треонин, аспартат, глутамат
Некоторые аминокислоты обнаружены в тканях организма, однако они не используются для построения белковых молекул (орнитин, цитруллин). Есть также аминокислоты (оксипролин и цистеин), которые образуются из других (пролин, цистин) после включения в состав белков.
К аминокислотам с разветвленной цепью относятся 3 (три) аминокислоты: валин, лейцин и изолейцин. Свое название аминокислоты с разветвленной цепью получили из-за структурных особенностей строения молекулы, что придает им ряд уникальных свойств. Помимо вхождения в структуру мышечной ткани (42% от общего количества незаменимых аминокислот) аминокислоты с разветвленной цепью играют первостепенную роль в обмене белков и снабжении мышц энергией. Основным источником энергии при интенсивных нагрузках служит гликоген печени и мышц, запасы которого быстро истощаются, и организм переходит к сжиганию свободных аминокислот - в первую очередь именно аминокислот с разветвленной цепью. В этом случае тренировка носит катаболический характер, то есть приводит не к росту, а к уменьшению мышц. Аминокислоты с разветвленной цепью активизируют обмен инсулина и стимулируют поступление других аминокислот в мышцы для последующего синтеза белка. Активизация обмена инсулина приводит к улучшению работы пищеварительных ферментов и метаболитов цикла Кребса (основного цикла обмена энергии в организме), способствующих восстановлению энергетического потенциала мышечных клеток. В мышцах аминокислоты с разветвленной цепью перерабатывают продукты обмена (молочную кислоту и др.) в аминокислоту аланин, которая участвует в образовании мышечного гликогена.
Функции
белков в клетках
живых организмов более разнообразны, чем функции других
биополимеров —
полисахаридов
и ДНК.
Так, белки-ферменты
катализируют протекание биохимических реакций и играют важную роль в
обмене веществ. Некоторые белки выполняют структурную или
механическую функцию, образуя цитоскелет,
поддерживающий форму клеток. Также белки играют ключевую роль в
сигнальных
системах клеток, при иммунном
ответе и в клеточном
цикле. С белками связано все многообразие
функций организма, однако наиболее важными из них являются:
каталитическая, транспортная, защитная, сократительная, структурная,
гормональная и питательная. Каталитическая функция белков
осуществляется с помощью специфических белков- катализаторов -
ферментов. При их участии увеличивается скорость различных реакций
обмена веществ и энергии в организме.
Транспортная функция белков
заключается в том, что при их участии происходит связывание и
доставка (транспорт) различных веществ от одного органа к другому.
Так, белок эритроцитов крови гемоглобин соединяется в легких с
кислородом, превращаясь в оксигемоглобин. Достигая с током крови
органов и тканей, оксигемоглобин расщепляется и отдает кислород,
необходимый для обеспечения окислительных процессов в тканях. Другие
белки крови связывают триглицериды, жирные кислоты, холестерин,
кальций, некоторые гормоны, витамины и другие вещества и
транспортируют их к месту использования или действия.
Защитную
функцию выполняют специфические белки (антитела), образующиеся в
организме. Они обеспечивают связывание и обезвреживание веществ,
поступающих в организм или появляющихся в результате
жизнедеятельности бактерий и вирусов. Защитную функцию выполняет
белок плазмы крови фибриноген, участвуя в свертывании крови и тем
самым уменьшая кровопотери.
Сократительную функцию выполняют
белки, в результате взаимодействиях которых происходит передвижение в
пространстве, сокращение и расслабление сердца, движение других
внутренних органов.
Структурная функция белков заключается в том,
что они составляют основу строения клетки; некоторые из них (коллаген
соединительной ткани, кератин волос, ногтей и кожи, эластин
сосудистой стенки и др.) выполняют почти исключительно структурную
функцию. В комплексе с липидами (преимущественно фосфолипидами) белки
участвуют в построении мембран клеток и внутриклеточных образований.
Гормональную функцию выполняют белки-регуляторы обмена веществ.
Они относятся к гормонам, которые образуются в железах внутренней
секреции, некоторых органах и тканях организма.
Питательная
функция осуществляется белками, которые являются резервными, или
питательными. Белки яйца обеспечивают рост и развитие плода, белки
молока служат источником питания для новорожденного.
Перечисленные
функции белков являются наиболее важными и специфичными, но ими не
ограничивается значение белков для жизнедеятельности организма.
Размер белка может измеряться в числе аминокислотных остатков или в дальтонах (молекулярная масса), но из-за относительно большой величины молекулы масса белка выражается в производных единицах — килодальтонах (кДа). Белки дрожжей, в среднем, состоят из 466 аминокислотных остатков и имеют молекулярную массу 53 кДа. Самый большой из известных в настоящее время белков — титин — является компонентом саркомеров мускулов; молекулярная масса его различных вариантов (изоформ) варьирует в интервале от 3000 до 3700 кДа. Титин камбаловидной мышцы (лат. soleus) человека состоит из 38 138 аминокислот[13].
Для определения молекулярной массы белков применяют такие методы, как гель-фильтрация, электрофорез в полиакриламидном геле, масс-спектрометрический анализ, седиментационный анализ и другие[14].
Размер белка может измеряться в числе аминокислотных остатков или в дальтонах (молекулярная масса), но из-за относительно большой величины молекулы масса белка выражается в производных единицах — килодальтонах (кДа). Белки дрожжей, в среднем, состоят из 466 аминокислотных остатков и имеют молекулярную массу 53 кДа. Самый большой из известных в настоящее время белков — титин — является компонентом саркомеров мускулов; молекулярная масса его различных вариантов (изоформ) варьирует в интервале от 3000 до 3700 кДа. Титин камбаловидной мышцы (лат. soleus) человека состоит из 38 138 аминокислот[13].
Для определения молекулярной массы белков применяют такие методы, как гель-фильтрация, электрофорез в полиакриламидном геле, масс-спектрометрический анализ, седиментационный анализ и другие[14].
Сравнительный размер молекул белков. Слева направо: антитело (IgG), гемоглобин, инсулин (гормон), аденилаткиназа (фермент) и глютаминсинтетаза (фермент)
Белки различаются по степени растворимости в воде. Водорастворимые белки называются альбуминами, к ним относятся белки крови и молока. К нерастворимым, или склеропротеинам, относятся, например, кератин (белок, из которого состоят волосы, шерсть млекопитающих, перья птиц и т. п.) и фиброин, который входит в состав шёлка и паутины[16]. Растворимость белка определяется не только его структурой, но внешними факторами, такими как природа растворителя, ионная сила и pH раствора[14].
Белки также делятся на гидрофильные и гидрофобные (водооталкивающие). К гидрофильным относится большинство белков цитоплазмы, ядра и межклеточного вещества, в том числе нерастворимые кератин и фиброин. К гидрофобным относится большинство белков, входящих в состав биологических мембран, — интегральных мембранных белков, которые взаимодействуют с гидрофобными липидами мембраны[17] (у этих белков, как правило, есть и гидрофильные участки).
Первичная структура — последовательность аминокислотных остатков в полипептидной цепи. Первичную структуру белка, как правило, описывают, используя однобуквенные или трёхбуквенные обозначения для аминокислотных остатков.
Генетический код
Генети́ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства генетического кода: триплетность (3 нуклеотида - 1 аминокислота), неперекрываемость (кодоны одного гена не перекрываются, хотя в настоящее время известны и перекрывающиеся гены <overlapping genes>), вырожденность (кодирование одной аминокислоты несколькими триплетами), однозначность (каждый кодон кодирует только одну аминокислоту), компактность (Г.к. не включает мелкие пробелы между кодонами - “запятые”), универсальность (хотя имеются и исключения).

В ДНК
используется четыре азотистых основания — аденин
(А),
гуанин (G),
цитозин (С),
тимин (T).
Буквы A
G C T
составляют алфавит
генетического кода. В
РНК используются те же
нуклеотиды, за исключением тимина,
который заменён похожим нуклеотидом — урацилом,
который обозначается буквой U
(А, Г,
Ц и Т
и У в русскоязычной
литературе). В молекулах ДНК
и РНК нуклеотиды
выстраиваются в цепочки и, таким образом, получаются
последовательности генетических букв. Каждую аминокислоту кодирует
комбинация из трёх нуклеотидов – триплет, или кодон. Кратко
путь переноса генетической информации обобщён в т.н. центральной
догме молекулярной биологии: ДНК ` РНК f белок.
В особых случаях
информация может переноситься от РНК к ДНК, но никогда не переносится
от белка к генам.
Реализация генетической информации в живых клетках (то есть синтез белка, кодируемого геном) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза мРНК на матрице ДНК) и трансляции генетического кода в аминокислотную последовательность (синтез полипептидной цепи на мРНК). Реализация генетической информации осуществляется в два этапа. В клеточном ядре на ДНК синтезируется информационная, или матричная, РНК (транскрипция). При этом нуклеотидная последовательность ДНК «переписывается» (перекодируется) в нуклеотидную последовательность мРНК. Затем мРНК переходит в цитоплазму, прикрепляется к рибосоме, и на ней, как на матрице, синтезируется полипептидная цепь белка (трансляция). Аминокислоты с помощью транспортной РНК присоединяются к строящейся цепи в последовательности, определяемой порядком нуклеотидов в мРНК. (Биосинтез)
Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. В 1953 году Уотсон и Крик опубликовали две работы: в первой говорилось о вторичной структуре ДНК[3], а во второй — о возможном механизме копирования ДНК путём матричного синтеза[4]. В последней работе, они указали на то, что определённая последовательность оснований является кодом, который несёт генетическую информацию. первым кто предложил абстрактную гипотезу кодирования, а также способ её проверки, был советский и американский физик-теоретик Георгий (Джордж) Гамов. В 1954 году Гамов опубликовал свою работу, в которой предложил в качестве механизма кодирования установление соответствия между боковыми цепями аминокислот и ромбовидными «дырами», образованными четырьмя нуклеотидами ДНК[6]. Позднее этот код был назван ромбическим или бубновым. Гамов предположил, что код может быть триплетным, а к 1965 году был установлен смысл всех 64 триплетов. Оказалось, что некоторые кодоны просто-напросто избыточны, то есть целый ряд аминокислот кодируется двумя, четырьмя или даже шестью триплетами.
| Первая | Вторая буква | Третья |
| «буква» |----------------------------------------------------------------------------------------------------------------------------------------------| «буква» |
|----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------|
| У | УУУ | Фенила- | УЦУ | Серин | УАУ | Тирозин | УГУ | Цистин | У |
|------------------------------------| ланин |------------| |------------| |------------| |---------------- |
| | УУЦ | | УЦЦ | | УАЦ | | УГЦ | | Ц |
|----------------------------------------------------------------------| |---------------------------------------------------------------------------------------|
| | УУА | Лейцин | УЦА | | УАА | Конец | УГА | Конец | А |
| | | | | | | синтеза | | синтеза | |
|------------------------------------| |------------| |------------| |-----------------------------------------------------|
| | УУГ* | | УЦГ | | УАГ | | УГГ | Триптофан | Г |
|----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------|
| Ц | ЦУУ | Лейцин | ЦЦУ | Пролин | ЦАУ | Гистидин | ЦГУ | Аргинин | У |
|------------------------------------| |------------| |------------| |------------| |---------------- |
| | ЦУЦ | | ЦЦЦ | | ЦАЦ | | ЦГЦ | | Ц |
|------------------------------------| |------------| |----------------------------------------------| |---------------- |
| | ЦУА | | ЦЦА | | ЦАА | Глутамин | ЦГА | | А |
|------------------------------------| |------------| |------------| |------------| |---------------- |
| | ЦУГ | | ЦЦГ | | ЦАГ | | ЦГГ | | Г |
|----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------|
| А | АУУ | Изолей- | АЦУ | Треонин | ААУ | Аспарагин | АГУ | Серин | У |
|------------------------------------| цин |------------| |------------| |------------| |---------------- |
| | АУЦ | | АЦЦ | | ААЦ | | АГЦ | | Ц |
|------------------------------------| |------------| |---------------------------------------------------------------------------------------|
| | АУА | | АЦА | | ААА | Лизин | АГА | Аргинин | А |
|----------------------------------------------------------------------| |------------| |------------| |---------------- |
| | АУГ* | Метионин | АЦГ | | ААГ | | АГГ | | Г |
|----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------|
| Г | ГУУ | Валин | ГЦУ | Аланин | ГАУ | Аспараги- | ГГУ | Глицин | У |
|------------------------------------| |------------| |------------| новая |------------| |---------------- |
| | ГУЦ | | ГЦЦ | | ГАЦ | кислота | ГГЦ | | Ц |
|------------------------------------| |------------| |----------------------------------------------| |---------------- |
| | ГУА | | ГЦА | | ГАА | Глутами- | ГГА | | А |
|------------------------------------| |------------| |------------| новая |------------| |---------------- |
| | ГУГ* | | ГЦГ | | ГАГ | кислота | ГГГ | | Г |
-----------------------------------------------------------------------------------------------------------------------------------------------------
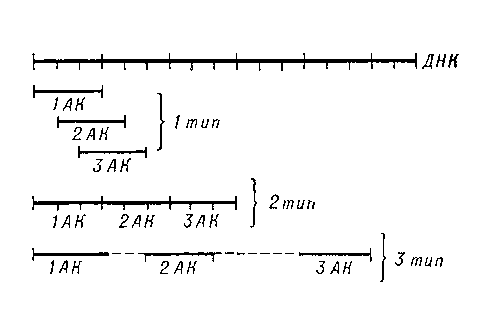
Большая советская энциклопедия. — М.: Советская энциклопедия. 1969—1978.
Биологические строительные блоки ядра клетки
Ядро клетки интересно, как и ее оболочка, тем, что его физко-химическая работа заключается в том, что оболочка ядра пропускает нужные вещества, подобно ситу, и отсеивает ненужные. Ядро клетки современные ученые считают самым важным в области лечения и научных разработок. Восстановление ядра — основной путь к восстановлению клетки. Поэтому особенно интересна структура ядерной оболочки, которая является живы фильтром и живым прибором, который отделяет живой материал ядра и пропускает нужные для существования ядра вещества.
Изоэлектрическая точка (радикалы)
Изоэлектрическая точка (pI) — кислотность среды (pH), при которой определённая молекула или поверхность не несёт электричекого заряда. Крепления по связи NH3 у молекулы имеют знак +, за счет лишнего атома водорода, а у СОО- не хватает атома водорода на конце. Амфотерные молекулы (цвиттер-ионы), или нейтральные молекулы содержат как положительные, так и отрицательные заряды. Заряд различных функциональных групп таких молекул может меняться в результате связывания или, наоборот, потери протонов H+. Это связано с тем, что водородные связи в молекуле и относительно легче изучать из-за того, что у водорода только один электрон и строение атома простейшее, а вычисление и техника оперируют меньшим количеством задач, которые нужно решить. И с тем, что водород имеет минимум сил притяжения (атом маленький, электрон один), а из-за этого связи водорода самые непрочные. При отрыве атома водорода на одном конце СООН получается СОО- , и, соответственно, отрицательный заряд. При NH3+ на этом конце один атом водорода тоже может оторваться, тогда NH2 имеет нейтральный заряд. У белковой молекулы, которая имеет на одном конце СОО-, а на другом NH3+ кислотность среды 7, у СОО-, а на другом NH2 заряд -, кислотность больше 7. У белков с СООН на одном конце и NH3+ на другом лишний протон дает положительный заряд и кислотность меньше 7. Нейтральные белки в магнитной среде и электрическом поле остаются на месте и никуда не движутся.
Величина изоэлектрической точки такой амфотерной молекулы определяется величинами констант диссоциаций кислотной и основной фракций:
pI = 0,5(рК1+рK2).
Растворимость амфотерных молекул, как правило, является минимальной при pH равной или близкой к изоэлектрической точке pI. Часто они в своей изоэлектрической точке выпадают в осадок. Аминокислоты и белки являются по своей природе амфотерными, так как содержат и кислотные, и осно́вные функциональные группы. Общий заряд белка определяется боковыми группами аминокислот, которые могут быть положительно- или отрицательно-заряженными, нейтральными или полярными.
Неполярные:аланин, валин, изолейцин, лейцин, пролин, метионин, фенилаланин, триптофан
Полярные незаряженные (заряды скомпенсированы) при pH=7: глицин, серин, треонин, цистеин, аспарагин, глутамин, тирозин
Полярные заряженные отрицательно при pH=7: аспартат, глутамат
Полярные заряженные положительно при pH=7: лизин, аргинин, гистидин
Общий заряд белка при pH ниже изоэлектрической точки является положительным. Наоборот, при pH выше изоэлектрической точки общий заряд белка — отрицательный. В самой изоэлектрической точке сумма положительных зарядов на белковой молекуле равна сумме отрицательных зарядов, поэтому будучи помещена в электрическое поле такая молекула не двигается.
Википедия. Статья Белки
Аминокислоты // Энциклопедический словарь Брокгауза и Ефрона: В 86 томах (82 т. и 4 доп.). — СПб., 1890—1907.
Эксперименты Миллера-Юри и обсуждения:
Miller S. L. Production of amino acids under possible primitive earth conditions. Science, v. 117, May 15, 1953
Miller S. L. and H. C. Urey. Organic compound synthesis on the primitive earth. Science, v. 130, July 31, 1959
Miller Stanley L. and Leslie E. Orgel. The origins of life on the earth. Englewood Cliffs, NJ, Prentice-Hall, 1974.
Общая биология. Учебник для 9 — 10 классов средней школы. Под ред. Ю. И. Полянского. Изд. 17-е, перераб. — М.: Просвещение, 1987. — 288с.
Аминокислоты, пептиды, белки. Под ред. Ю. В. Митина
Научно-технический энциклопедический словарь. Статья: Генетический код. http://dic.academic.ru/dic.nsf/ntes/1006/%D0%93%D0%95%D0%9D%D0%95%D0%A2%D0%98%D0%A7%D0%95%D0%A1%D0%9A%D0%98%D0%99
Биологический энциклопедический словарь. Статья: Генентический код. (Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986.)
.(Источник: «Биология. Современная иллюстрированная энциклопедия.» Гл. ред. А. П. Горкин; М.: Росмэн, 2006.) http://dic.academic.ru/dic.nsf/dic_biology/1151/%D0%93%D0%95%D0%9D%D0%95%D0%A2%D0%98%D0%A7%D0%95%D0%A1%D0%9A%D0%98%D0%99
(Источник: «Англо-русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд-во ВНИРО, 1995 г.)
Большая советская энциклопедия. — М.: Советская энциклопедия. 1969—1978.
Известные ученые, история открытий:
ДАЛЬТОН (Долтон) (Dalton) Джон (6 сентября 1766-1844) - английский химик и физик, создатель химического атомизма. Установил (1803) закон кратных отношений, ввел понятие «атомный вес», первым определил атомные веса (массы) ряда элементов. Открыл газовые законы, названные его именем. Первым (1794) описал дефект зрения, которым страдал сам, позже названный дальтонизмом.
ГЕЙ-ЛЮССАК (Gay-Lussac) Жозеф Луи (1778-1850) - французский химик и физик, иностранный почетный член Петербургской АН (1829). Открыл газовые законы, названные его именем. Открыл бор (1808, совместно с Л. Тенаром). Получил (1811) безводную синильную кислоту и исследовал (1815) ее количественный состав, открыл (1815) дициан. Построил первые диаграммы растворимости (1819). Усовершенствовал методы элементного и объемного химического анализа, технологию производства серной кислоты (башня Гей-Люссака). Совместно с М. Шеврелем получил (1825) патент на изготовление стеариновых свечей.
БРАКОННО (Braconnot) Анри (1780-1855) - французский химик. Труды по химии природных соединений. Получил (1819) виноградный сахар (глюкозу) гидролизом целлюлозы. Выделил (1820) из гидролизата белка аминокислоты глицин и лейцин. Синтезировал (1833) тринитратцеллюлозу — один из первых полимерных материалов.
ШТРЕККЕР (Strecker) Адольф (1822-1871) - немецкий химик-органик, труды по исследованию и синтезу природных соединений. Открыл названные его именем реакции, в т. ч. получение альфа-аминокислот из аммиака (1850).
ФИШЕР Эмиль Герман (1852-1919) - немецкий химик-органик, создатель научной школы, основоположник химии природных соединений, иностранный член-корреспондент (1899) и иностранный почетный член (1913) Петербургской АН. Исследовал строение и синтезировал ряд производных пурина: кофеин, гуанин, аденин и др. Ввел номенклатуру, создал рациональную классификацию и осуществил синтез многих углеводов. Открыл специфичность действия ферментов. Основополагающие исследования по химии белков. Нобелевская премия (1902).
КОССЕЛЬ (Kossel) Альбрехт (1853-1927) Биохимик. Труды по химии ядерных белков (протаминов и гистонов), нуклеиновых кислот и нуклеопротеидов. Создал одну из первых теорий строения белков. Нобелевская премия (1910).
ЗЕЛИНСКИЙ Николай Дмитриевич [6 февраля (25 января) 1861, Тирасполь Херсонской губернии, ныне Молдавия — 31 июля 1953, Москва], российский химик-органик, автор фундаментальных открытий в области синтеза углеводородов, органического катализа, каталитического крекинга нефти, гидролиза белков и противохимической защиты.
ДАНИЛЕВСКИЙ Александр Яковлевич (1838-1923) - российский биохимик, основатель первой русской научной школы. Брат В. Я. Данилевского. Основные труды по химическому строению и обмену белков, энзимологии, биохимии питания.
БУТЛЕРОВ Александр Михайлович (1828-1886) - российский химик-органик, академик Петербургской АН (1874). Создал (1861) и обосновал теорию химического строения, согласно которой свойства веществ определяются порядком связей атомов в молекулах и их взаимным влиянием. Первым объяснил (1864) явление изомерии. Открыл полимеризацию изобутилена. Синтезировал ряд органических соединений (уротропин, полимер формальдегида и др.). Труды по сельскому хозяйству, пчеловодству. Поборник высшего образования для женщин.
АБДЕРГАЛЬДЕН (Abderhalden) Эмиль (1877-1950) - швейцарский биохимик, иностранный член-корреспондент АН СССР (1925). В 1904-45 работал в Германии. Исследовал структуру и функции белков. Синтезировал (совместно с Э. Г. Фишером) полипептид из 19 аминокислот (1916).
Определение аминокислотной последовательности первого белка — инсулина — методом секвенирования белков принесло Фредерику Сенгеру Нобелевскую премию по химии в 1958 году.
Первые трёхмерные структуры белков гемоглобина и миоглобина были получены методом дифракции рентгеновских лучей, соответственно, Максом Перуцем и Джоном Кендрю в конце 1950-х годов[2][3], за что в 1962 году они получили Нобелевскую премию по химии.